Chapitre 1 : L’origine du génotype des individus
Diaporama chap 1Révision : TD la conservation des génomes
Term spé SVT. Test 1. Génétique.
Rappels
Term spé SVT. Test 2.
Génétique. La notion de clone
Term spé SVT. Test 3.
Génétique. La lignée clonale.
Term spé SVT. Test 4.
Génétique. Les lois de Mendel.
Term spé SVT. Test 5.
Génétique. Méiose et brassage
Term spé SVT. Test 6.
Génétique. Lien entre les gènes.
Term spé SVT. Test 7.
Génétique. Maladies génétiques
Term spé SVT. Test 8.
Génétique. Les accidents
génétiques.
Term spé SVT. Test 9.
Génétique. Les familles
multigéniques.
Plusieurs activités peuvent être réalisées en ligne pour réactiver certaines notions de spécialité première :
- Réviser les étapes de la méiose
- Revoir comment on obtient un caryotype
- Réviser les états des chromosomes au cours du cycle cellulaire
Comment les divisions et la
fécondation participent-elles à
l’émergence de nouveaux génomes ?
I La
conservation
des génomes : stabilité génétique et
évolution clonale
A La mitose , la
méiose, la fécondation et la stabilité
génétique au sein de l’espèce
Les organismes
pluricellulaires
présentant une reproduction sexuée évoluent selon
des cycles, où une phase
haploïde (un seul exemplaire de chaque chromosome : n) et une
phase diploïde
(deux exemplaires de chaque chromosome 2 n avec n le nombre de paire)
alternent.
La reproduction
sexuée comprend
toujours deux phénomènes fondamentaux : la
méiose et la fécondation :
- la méiose fait passer le
nombre de chromosome par cellule de 2 n à n
- la fécondation en réunissant deux gamètes haploïdes (n), un spermatozoïde et un ovule à n chromosomes, reconstitue les paires de chromosomes homologues, rétablit la diploïdie et conduit à une cellule œuf (zygote), cellule diploïde. Un individu est constitué de cellules diploïdes (2n) qui résultent de mitoses successives à partir d’une cellule œuf initiale .
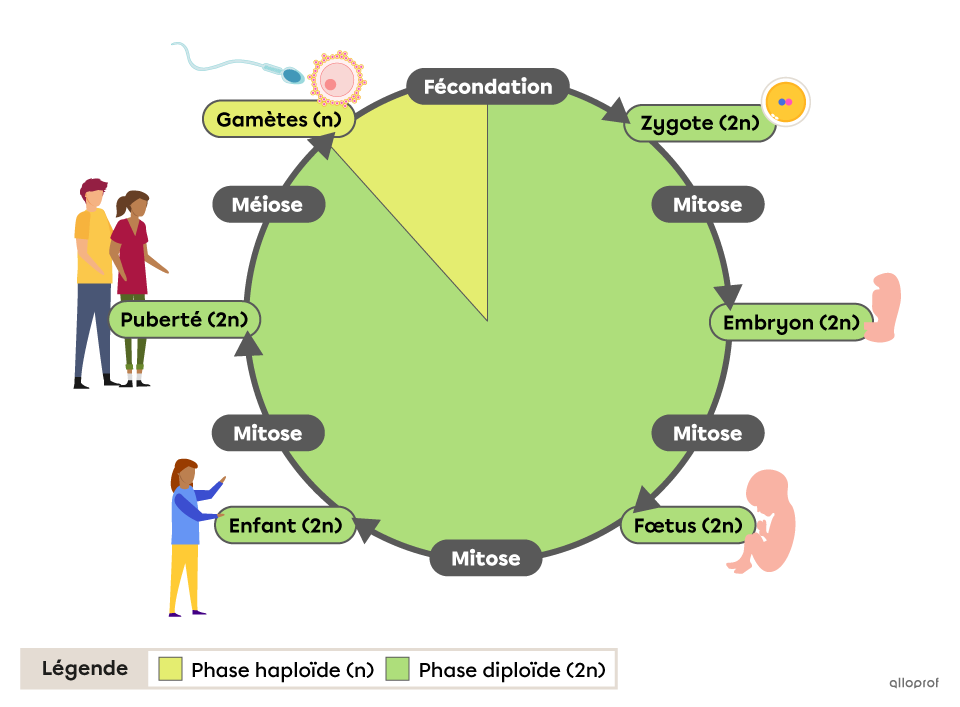
Ces mitoses
successives permettent d’obtenir un ensemble de cellules en
théorie
génétiquement identique, c’est-à-dire un
clone, puisque la mitose est précédée
d’un mécanisme efficace de copie de l’information
génétique : la réplication de
l’ADN.
La mitose est une reproduction conforme : elle conserve le caryotype de la cellule mère ainsi que l’information génétique. Autrement dit, toutes les cellules issues des mitoses successives d’une cellule mère possèdent la même information génétique aux mutations près : elles constituent un clone.

Variation de la quantité
d’ADN
dans le noyau d’une cellule somatique au cours d’un cycle
mitotique.
Les individus issus de la reproduction sexuée, ressemblent à leurs parents, à leurs frères et sœurs mais sont génétiquement uniques (pas les mêmes allèles de gènes). Ainsi, si la reproduction sexuée assure la stabilité de l’espèce en maintenant le caryotype c’est-à-dire la totalité des gènes de l’espèce, elle est aussi source de variabilité génétique des individus à l’intérieur de l’espèce.
La diversité
éclairée par la génétique
https://www.science-et-vie.com/article-magazine/la-diversite-eclairee-par-la-genetique

TD 12 l'organisme : une multitude de clone cellulaire
CorrectionB
L’évolution
clonale et la diversité génétique au sein
d’un clone
Chaque individu est constitué d’une mosaïque de clones présentant de faibles variations génétiques liées à ces mutations accumulées.
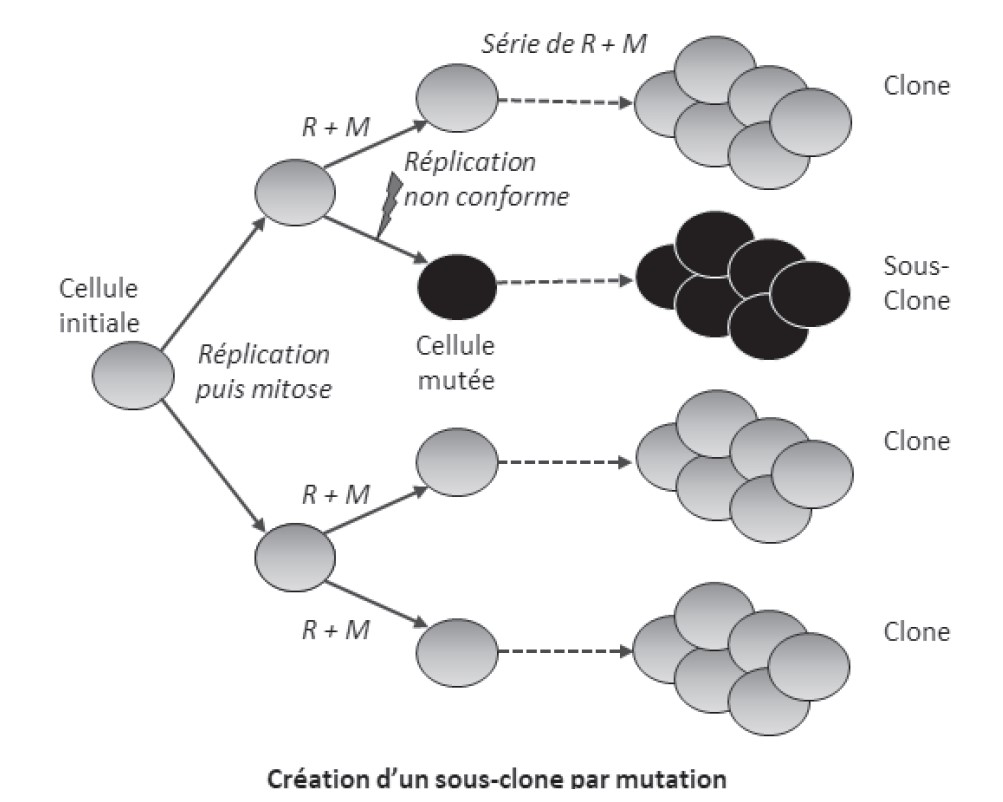
- Les
mutations affectant une cellule deviennent pérennes pour toute
la lignée cellulaire qui dérive du mutant, formant ainsi
un sous clone particulier.
En l’absence d’échanges génétiques avec l’extérieur, la diversité génétique dans un clone résulte de l’accumulation de mutations successives dans les différentes cellules.

Tout accident génétique irréversible (perte de gène par exemple) devient pérenne pour toute la lignée (sous-clone) qui dérive du mutant. Ces mutations ne sont transmises à la génération suivante que si elles affectent les cellules germinales.
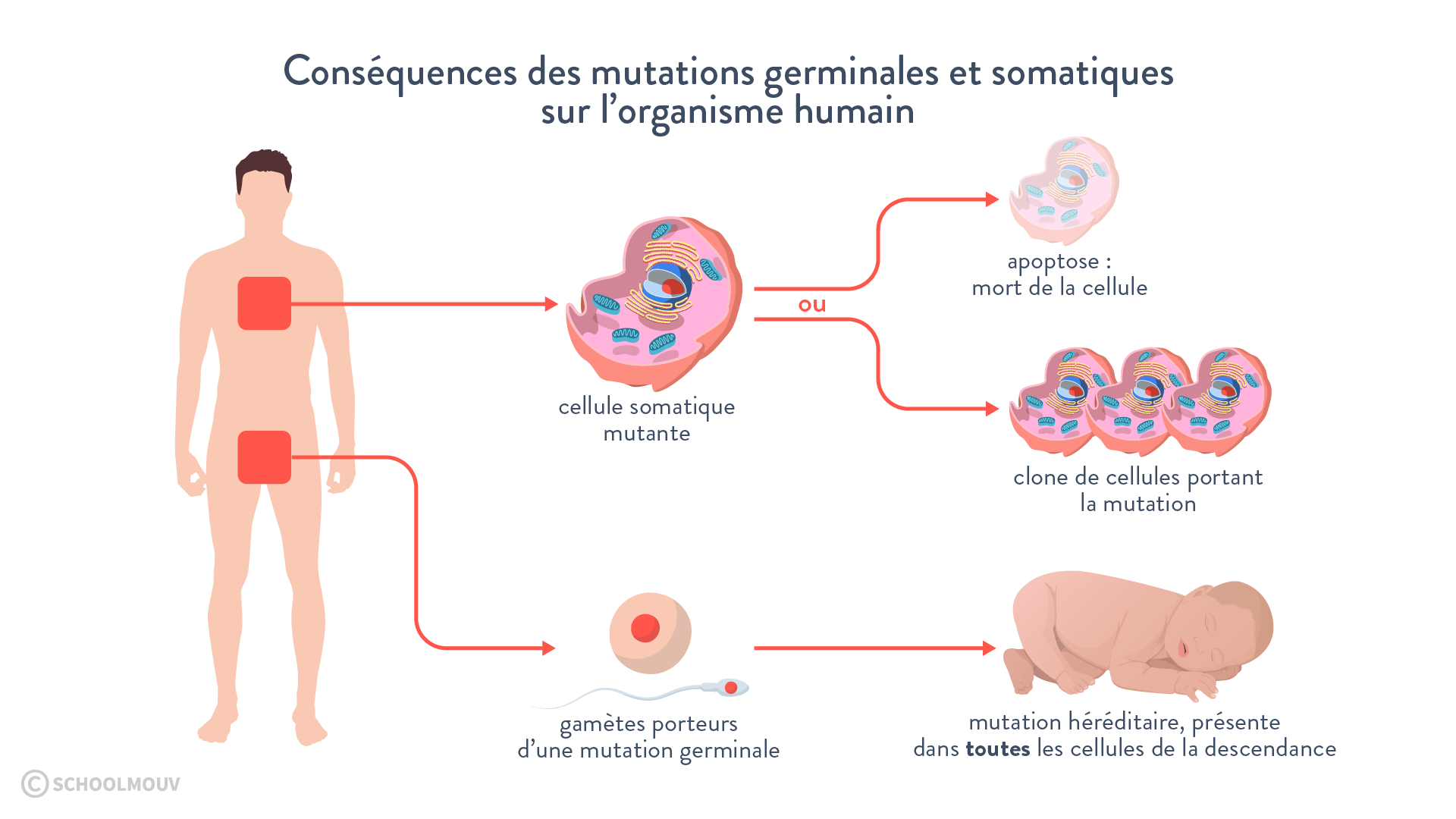

II Le brassage des
génomes à chaque génération
Comment la
méiose peut-elle produire des gamètes
génétiquement diversifiés ?
A La méiose assure un brassage interchromosomique
TP 13 Comprendre les lois de Mendel (version 1)TP 13 Comprendre les lois de Mendel (version 2)
Activité 1 comprendre les lois de Mendel
Prise de notes des éléments importants de la vidéo pour identifier la démarche scientifique mise en place par Mendel pour comprendre la transmission des caractères d’une génération à l’autre.
Mendel et ses petits-pois, le début de la génétique.
Sachant que le caractère lisse ou ridé des grains de pois est gouverné par un seul gène et deux allèles, proposer une interprétation chromosomique des résultats décrits ci-dessous en F1 et F2. Pour cela, écrire les génotypes des différents individus.
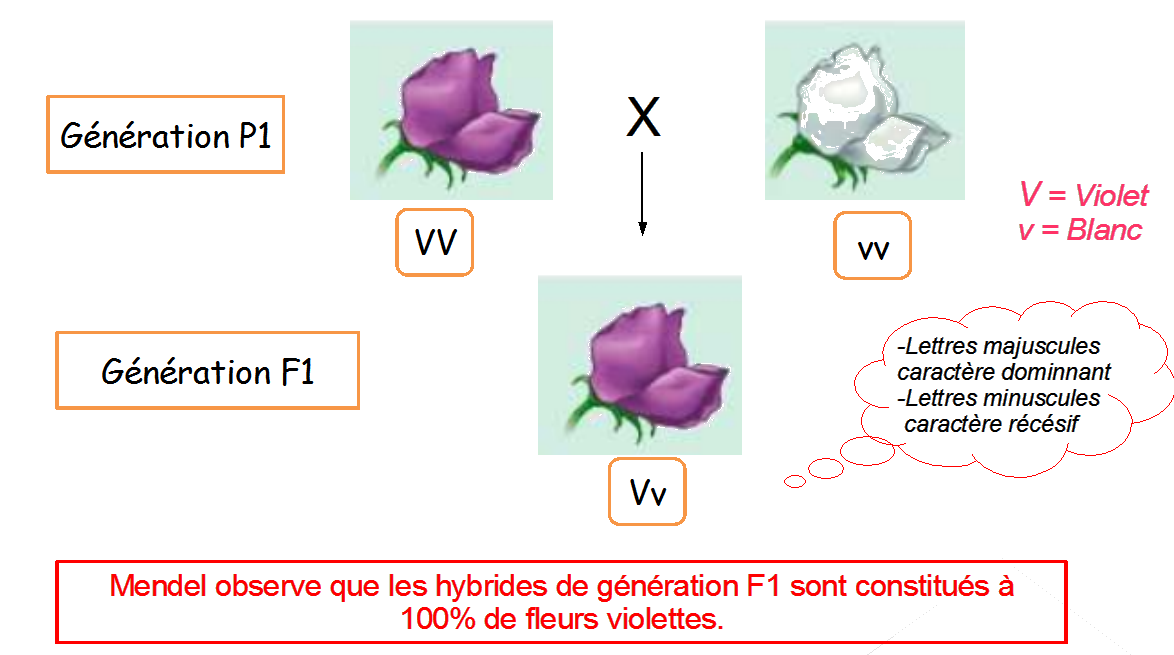
Première loi : Loi d’uniformité des hybrides de première génération (tous les hybrides de F1 sont 100% identiques)
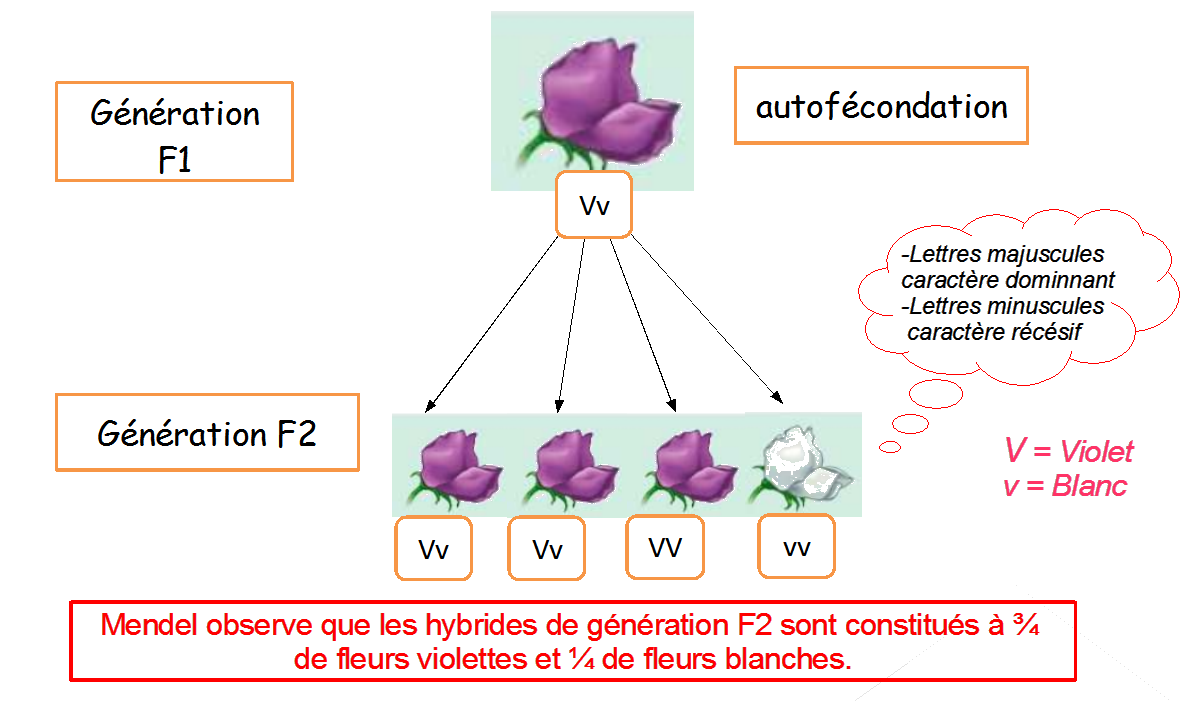
Deuxième loi : Loi de disjonction des allèles (1 caractère ne s’étant pas exprimé lors de la 1ère génération peut s’exprimer à la seconde génération)

Troisième loi : indépendance de la transmission des caractères (les 2 premières lois sont valables même avec plusieurs caractères
TP 14 le brassage des gènes lors de la reproduction
La drosophile (mouche du vinaigre) est un matériel biologique intéressant : son élevage est aisé, sa reproduction est rapide et sa garniture chromosomique est très simple (2n = 8 chromosomes). Elle possède de nombreuses mutations connues et localisées sur les chromosomes, dont la transmission est facile à observer (longueur des ailes, couleur du corps, couleur des yeux, position des organes…). On peut aussi observer un dimorphisme sexuel entre les mâles et les femelles : les femelles sont plus grandes que les mâles, l’abdomen de la femelle est de forme pointue, avec des segments terminaux de couleur claire, l’abdomen du mâle est plus arrondi, avec des segments terminaux très foncés. Cependant, l’analyse génétique chez les diploïdes est plus difficile que chez les haploïdes car :
- les gamètes issus de la méiose ne sont pas classés ;
- chaque gène s’exprime avec deux
allèles, il y a donc des effets de dominance et de
récessivité dans le phénotype.
L’observation des phénotypes ne permet donc pas d’en déterminer directement le génotype, il faudra trouver un autre moyen pour déterminer le génotype d’une drosophile.
On appellera F1 la première génération issue du croisement de deux parents P1 et P2, et F2 la deuxième génération issue du croisement de deux individus F1.
Comprendre le brassage sous forme d’animation.
Matériels :
- Lame présentant les phénotypes parentaux ;
- Lame présentant les résultats de la F1 ;
- Plaques présentant les résultats du croisement test : plaque « A » ou « E » ;
- Loupe binoculaire équipée d’une caméra ;
- Loupe à main ;
- Logiciel « Mesurim 2 » et sa fiche méthodologique
Les conséquences génétiques de la fécondation sous forme d’échiquier de croisement.
Des perturbations dans la répartition des chromosomes au cours de la méiose conduisent à des anomalies du nombre de chromosomes. Ces anomalies peuvent se produire au cours de chaque division.
- TP Un modèle génétique du brassage génétique : Sordaria & périthèces de Sordaria
- TP Absence de crossing-over chez les drosophiles mâles & plaques b/vg
- TP La couleur noire du corps chez la drosophile & plaques b/vg et eb/vg
- TP Crossing-over chez la drosophile & plaque b/vg
Chez les mammifères la
méiose se déroule dans les organes de reproduction
(testicules et ovaires) et
aboutit à la formation des gamètes.
Dans tous les cas la
méiose est :
- toujours
précédée d’une phase de réplication
semi conservative de l’ADN qui forme 2 chromatides identiques de
chaque chromosome,
- se compose de deux divisions cellulaires successives.
a-
La première division de méiose est une réduction
chromatique
La première
division
est dite réductionnelle, elle permet le passage de 2 n à
n avec séparation des
chromosomes homologues de chaque paire.
b-
La deuxième division de méiose est une séparation
des chromatides
La 2eme division de méiose est équationnelle, c’est à dire que le nombre de chromosomes se maintient, seules les chromatides de chaque chromosome se séparent (anaphase II), c’est pour cela qu’elle est comparable à une mitose.

Elle
présente également 4 phases, dont l’anaphase
2 qui permet la séparation des
chromatides par rupture des centromères. A l’issue de
cette deuxième division,
4 cellules haploïdes sont formées.
Les individus issus de la reproduction sexuée, ressemblent à leurs parents, à leurs frères et sœurs mais sont génétiquement uniques (pas les mêmes allèles de gènes). Ainsi, si la reproduction sexuée assure la stabilité de l’espèce en maintenant le caryotype c’est-à-dire la totalité des gènes de l’espèce, elle est aussi source de variabilité génétique des individus à l’intérieur de l’espèce.
TP 15 le brassage des gènes lors de la reproduction
B la méiose
réalise un brassage intrachromosomique
Lors de la prophase 1 de la méiose les chromosomes homologues de chaque paire sont étroitement appariés. On observe en effet des enjambements entre leurs chromatides qui se croisent en formant des chiasmas aussi appelés des crossing over.

A
leur niveau se produisent des échanges des portions de
chromatides qui
aboutissent à des échanges d’allèles du
même gène. On dit qu’il y a eu recombinaison
homologue et formation de chromatides recombinées
différentes de celles des
parents appelées chromatides parentales. Du coup, les
chromosomes ne
contiennent plus la même information génétique que
ceux d’origine. Les
emplacements des ces échanges varient d’une méiose
à une autre et sont
aléatoires, ce qui entraîne une variabilité des
résultats de ce brassage.
A la fin de la
prophase 1 de la méiose certains chromosomes n’ont pas les
2 chromatides
identiques, ils n’ont donc pas les mêmes allèles sur
leurs deux chromatides.
Les descendants de première génération (P1 x P2 = F1) permettent de déterminer la dominance ou la récessivité des allèles. 100% de phénotypes identiques confirment que les parents étaient de lignée pure.
| A | 100 % des F1 sont de génotype (A//a) et de phénotype [A] | |
| a | (A//a) |
Le
croisement test ou backcross (F1 x double
récessif) permet
de trouver le génotype des gamètes produits par
l’individu de la F1. En effet,
les gamètes du double récessifs n’apportent que des
allèles récessifs, le
phénotype des individus obtenus est donc imposé par les
allèles des gamètes de
l’individu F1.

Les phénotypes des
individus issus du test cross représentent les génotypes
des gamètes de
l’individu F1.
Les différentes
proportions obtenues à l’issu de ces croisements test
apportent des
informations supplémentaires :
- Pour un caractère donné, si le
résultat du test cross donne 50%- 50% c’est que le
caractère est sous la dépendance d’un seul
gène pour la détermination du phénotype
A a a (A//a) (a//a) [A] 50 % [a] 50 % - Pour un caractère donné, si le
résultat du test cross donne des pourcentages inégaux,
c’est que deux ou plusieurs gènes sont impliqués
dans l’établissement de ce caractère unique.
- S’il y a 4 types de gamètes dans des proportions équiprobables, c’est que les deux gènes ont été brassés de manière indépendante, ils sont donc sur des chromosomes différents, on dit que les deux gènes sont indépendants. Quand on obtient 4 types de gamètes équiprobables, on a mis en évidence un brassage interchromosomique qui a lieu en anaphase I de méiose.
-
A ; B A ; b a ; B
a ; b
a ; b (A//a ; B//b) (A//a ; b//b) (a//a ; B//b) (a//a ; b//b) [AB] 25 % [Ab] 25 % [aB] 25 % [ab] 25 %
- S’il y a 4 types de gamètes avec des proportions différentes, c’est que les deux gènes n’ont pas été brassés de manière indépendante. Certains gamètes ont une plus faible probabilité de se former, ils nécessitent la réalisation d’un événement peu probable, le crossing-over, entre les deux gènes. C’est que les deux gènes sont sur la même paire de chromosomes, on dit que les 2 gènes sont liés.
-
AB Ab aB
ab
ab (AB//ab) (Ab//ab) (aB//ab) (ab//ab) [AB]> 25 % [Ab] < 25 % [aB]< 25 % [ab] > 25 %
Quand on obtient 4 types de gamètes non
équiprobables, on a
mis en évidence un brassage interchromosomique en Anaphase 1 et
un brassage
intrachromosomique en Prophase I.
Remarque
1 : Dans le cas des
croisements de deux individus différant par deux
caractères, il faut considérer
d’abord chacun des caractères indépendamment de
l’autre afin de déterminer si
chaque caractère est contrôlé par un seul ou
plusieurs gènes. Dans le cas où
deux gènes sont impliqués pour un de ces deux
caractères, il faut déterminer
s’ils sont liés ou indépendants
Remarque
2 : Chez les drosophiles
le mâle ne fait jamais de crossing-over, c’est pour
cela que l’on
choisit toujours le mâle récessif pour le test cross.
C La fécondation
amplifie le brassage génétique
La
fécondation
ajoute un brassage supplémentaire car elle mélange deux
lots de chromosomes
venant de deux êtres différents. Ces 2 individus
produisent des gamètes de
génotype variable qui sont réunis au hasard lors de
la fécondation qui
est ainsi source de variabilité. La fécondation
réunit deux gamètes au
hasard et reconstitue les couples d’allèles.
Si
l’on ne considère
que le seul brassage interchromosomique, le nombre de
cellules-œufs différentes
que la fécondation peut engendrer est 2²³ x
2²³ = 8 388 608 x 8 388 608 =
70 368 744 177 664 soit plus de 70 milliers de milliards de
combinaisons
chromosomiques, donc aucune chance pour que 2 personnes aient
exactement le
même génome (et ces calculs ne tiennent pas compte des
crossing-over) ! Le
nombre de combinaisons génétiques possibles dans les
gamètes est d’autant plus
élevé que le nombre de gènes à
l’état hétérozygote est plus grand chez les
parents.
La fécondation en
réunissant au hasard un gamète mâle et un
gamète femelle, amplifie donc
considérablement le brassage génétique.
La méiose et la
fécondation réalisent un brassage génétique
qui assure l’unicité des
descendants.
III
Transmission
d’un caractère et phénotypes associés
Dans le cas de
l’espèce humaine,
l’étude de la transmission des allèles repose sur
des analyses généalogiques
familiales mais aussi sur des données issus des techniques
modernes
d’exploration de l’ADN : séquençage, PCR et
analyses biostatistiques.
Comment
la combinaison de deux allèles détermine-t-elle le
phénotype ? Comment
l’exploitation des informations génétiques et
généalogiques permet-elle de
prédire ou de suivre la transmission des allèles ?
A
L’examen des
arbres généalogiques permet de déterminer les
modalités de transmission d’un
allèle muté
L’étude
statistique
sur une seule famille ne présente pas
d’intérêt dans l’espèce humaine.
L’analyse
génétique peut se fonder sur l’étude de la
transmission héréditaire des
caractères observables (phénotype) dans des croisements
issus le plus souvent
de lignées pures (homozygotes) et ne différant que par un
nombre limité de
caractères. Dans le cas de l’espèce humaine,
l’identification des allèles
portés par un individu s’appuie d’abord sur une
étude au sein de la famille, en
appliquant les principes de transmission héréditaire des
caractères :
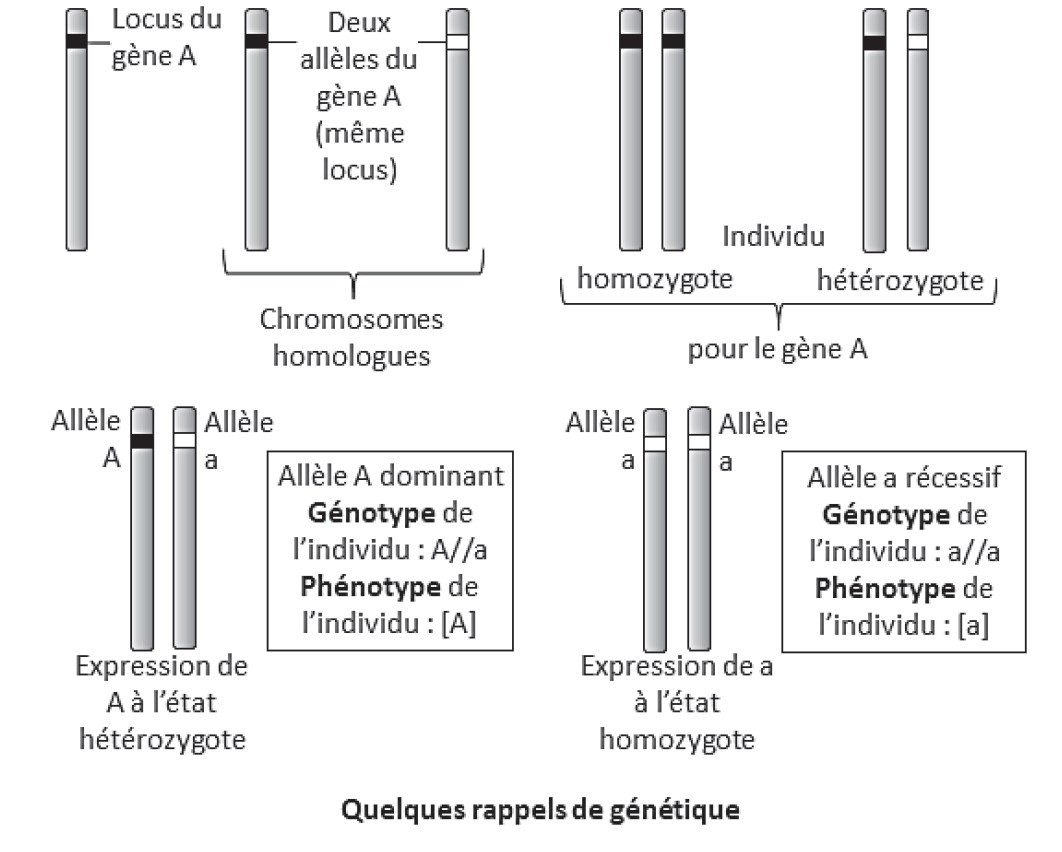
- le caractère dominant ou récessif d’une maladie la position du gène impliqué sur les chromosomes (autosomes ou gonosome)

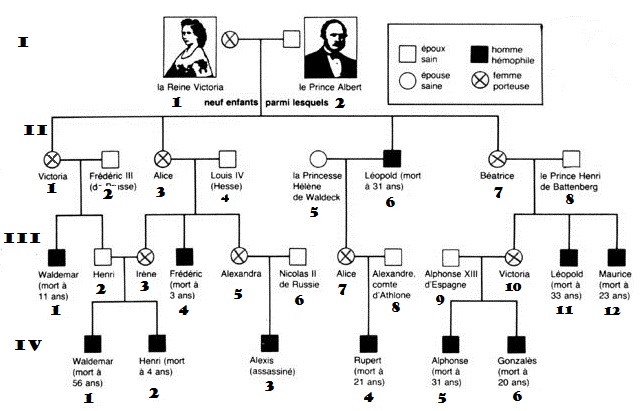
Un exemple très classique, l'hémophilie
- le risque génétique. Le caractère hémophile est transmis de la part des parents à la progéniture de manière mendélienne. Ce motif peut être analysé à l’aide d’un carré de Punnett. (https://fr.wikihow.com/travailler-avec-des-carr%C3%A9s-de-Punnett)
- (https://www.alloprof.qc.ca/fr/eleves/bv/sciences/les-croisements-les-lois-de-mendel-s1235)
Un autre exemple très classique, le daltonisme (voir vidéo de DTC qui répondra aux questions habituelles)
|
Carré de Punnett |
Parent 1 |
||
|
Parent 2 |
Gamètes |
S / |
s/ |
|
S / |
S//S |
S//s |
|
|
s / |
s//S |
s//s |
|
B Des
techniques de séquençage de l’ADN et de
bio-informatique
donnent directement accès au génotype
Nous parlons beaucoup
dans ce cours de séquences génomiques ou séquences
d’ADN, que nous voyons pour
des raisons algorithmiques sous forme de chaînes de
caractères.
Comment ces séquences, ces
chaînes de caractères, sont-elles
obtenues ?
- techniques modernes d’exploration
de l’ADN : le séquençage
Le
séquençage de l’ADN
est une technique qui a révolutionné la biologie
moléculaire dans les années
1970. La connaissance de sa séquence,
c’est-à-dire la succession des bases
de l’ADN , est aujourd’hui de plus en plus facile à
déterminer. Les appareils
qui servent à mener cette opération de
séquençage sont appelés séquenceurs.
Le mécanisme
du séquençage de l’ADN est le suivant :
- Dans un tube à essai, est placé
l’ADN à séquencer, des nucléotides
(A,T,G,C), une amorce, des didésoxynucléotides en
petite quantité et de l’ADN polymérase. Chaque
didésoxynucléotide est marqué par un fluorochrome
différent (A vert, T rouge, G jaune et C bleu), une chaîne
qui se termine par exemple par un A sera verte.
Séquence à
séquencer :
|
A |
C |
A |
G |
A |
C |
G |
A |
G |
G |
G |
C |
C |
G |
T |
A |
G |
G |
Commence la synthèse
du brin complémentaire de l’ADN à séquencer
par l’ADN polymérase.
- Lorsque
l’ADN polymérase choisit par hasard un
didésoxynucléotide (ce qui est rare puisqu’il y en
a moins que des nucléotides) et qu’elle
l’incorpore dans la chaîne en synthèse, celle-ci
s’interrompt prématurément.
- On
obtient ainsi des chaînes de toutes les tailles (correspondant
à un arrêt de la synthèse à chaque
nucléotide) et beaucoup de fragments d’une même
taille.
- On sépare ainsi par
électrophorèse les chaînes d’ADN obtenues en
fonction de leur taille. Plus les chaînes sont courtes, plus
elles migrent loin et tous les fragments d’une même taille
migrent à la même distance. On obtient alors une
succession de bandes colorées, chacune correspondant au dernier
nucléotide incorporé. Il suffit alors de lire la
succession des couleurs pour connaître l’ordre des
nucléotides, c’est-à-dire la séquence de
l’ADN, étape assurée automatiquement par les
détecteurs du séquenceur.
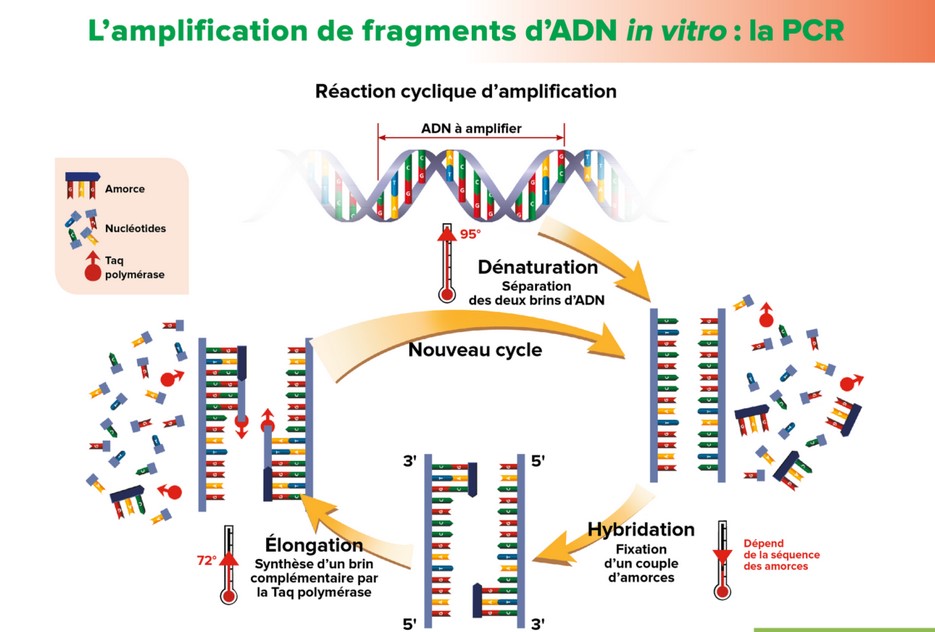
- techniques modernes
d’exploration de l’ADN : la PCR (voir TP)
L’amplification en chaîne par polymérase (Polymerase Chain Reaction en anglais) est une méthode de biologie moléculaire d’amplification génique in vitro. Elle permet de dupliquer en grand nombre (avec un facteur de multiplication de l’ordre du milliard) une séquence d’ADN ou d’ARN connue, à partir d’une faible quantité (de l’ordre de quelques picogrammes) d’acide nucléique et d’amorces spécifiques constituées d’oligonucléotides de synthèse de 20 à 25 nucléotides.
- techniques modernes
d’exploration de l’ADN : les analyses biostatistiques.
Les progrès de la
bioinformatique donnent directement accès au génotype de
chaque individu comme
à ceux de ces ascendants et descendants. Une collection
annotée de toutes
les séquences d’ADN disponibles publiquement est
déposée directement par les
laboratoires (GenBank
au NCBI , DNA
DataBank of Japan
(DDBJ), European Molecular Biology Laboratory (EMBL).
L’utilisation de bases
de données informatisées permet d’identifier des
associations entre certains
gènes mutés et certains phénotypes.Le
développement des techniques de
séquençage de l’ADN et les progrès de la
bio-informatique donnent directement
accès au génotype des individus. Grâce aux
techniques de séquençage, il est
possible d’établir les séquences des allèles
de certains gènes en particulier
les gènes responsables de maladies.
L’utilisation de bases de données informatisées permet d’identifier des associations entre certains gènes mutés et certains phénotypes. BLAST est une méthode de recherche heuristique utilisée en bioinformatique. Il permet de trouver les régions similaires entre deux ou plusieurs séquences de nucléotides ou d’acides aminés, et de réaliser un alignement de ces régions homologues.
(How to Use BLAST for Finding and Aligning DNA or Protein Sequences)
IV Les accidents
génétiques de la méiose
Comment les anomalies chromosomiques contribuent-elles à la diversité des génomes et à l’évolution des espèces ?
TD 16 Des accidents lors de la diversification des génomes
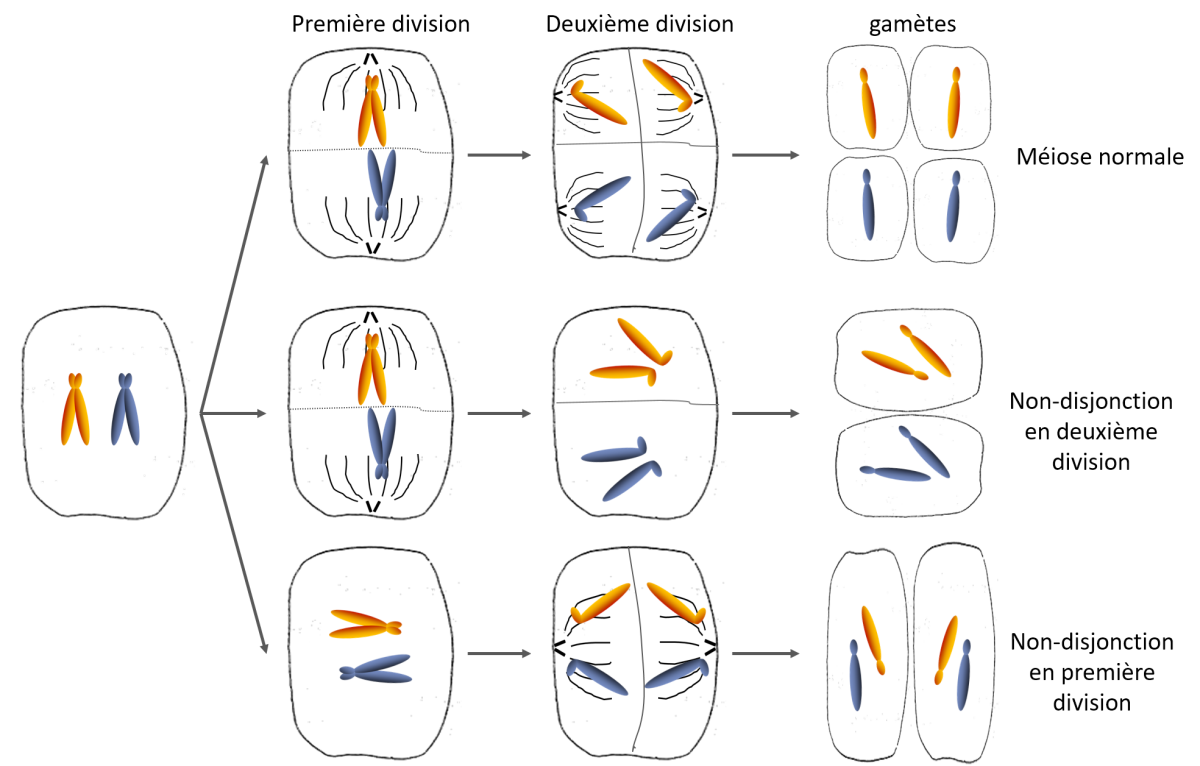
CorrectionA Migration
anormale des chromosomes au cours de la méiose
a Anomalie
de disjonction chromosomique
Des perturbations dans la
répartition des chromosomes au cours de la méiose
conduisent à des anomalies du
nombre de chromosomes. Ces anomalies peuvent se produire au cours de
chaque
division :
- soit en anaphase 1 une
paire d’homologue ne se sépare pas,
- soit en anaphase 2 :
un chromosome ne sépare pas ses 2 chromatides, elles se
séparent après.
(Caryotype humain à 2n =45 :
individus monosomiques ou 2n= 47 : chromosomes individus trisomiques)

b Anomalie
de cytodiérèse
Une
absence de cytodiérèse à la télophase
de la 2eme division méiotique peut
entraîner la formation de 2 gamètes au lieu de 4.
L’hybride résultant des
gamètes de ce type serait directement tétraploïde et
fertile (les chromosomes
qui s’ajoutent viennent du même organisme =
autopolyploïdie).
L’autopolyploïdisation
est entre autres le mécanisme à l’origine du
blé cultivé appelé Blé tendre :
Hervé Le Guyader l’explique dans cet article de Pour la
Science de Décembre
2018 "Comment le blé est devenu tendre ?"
B Des crossing
over inégaux à l’origine de duplication de
gènes
La possibilité de
survenue d’anomalies lors du déroulement de la
méiose (crossing-over inégal)
implique la possible duplication de certains gènes. De nouveaux
gènes peuvent
apparaître par mutations du gène préexistant.
Schéma
à l’origine de la duplication :

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Une fois
dupliquée, la copie obtenue s’insère sur le
même chromosome ou sur un autre
(sur un autre locus). C’est la transposition. Les
différences entre
les copies s’expliquent par l’accumulation de mutations
indépendantes après les
duplications, les deux versions du gène dupliqué,
d’abord identiques, deviennent
de plus en plus différentes.
Les innovations
génétiques sont aléatoires et leur nature ne
dépend pas des caractéristiques du
milieu.
Ces accidents,
souvent létaux, engendrent parfois une
diversification importante des génomes
et jouent un rôle essentiel dans l’évolution
biologique :
- Il existe des familles multigéniques.
Pour ces familles, il existe des similitudes de séquences et
elles sont classiquement interprétées comme un indice de
parenté, de sorte que les gènes d’une même
famille sont considérés comme dérivant tous
d’un gène ancestral commun.

Schéma
représentant l’origine des différents gènes
d’une même famille multigénique (Les opsines)
- il apparaît des barrières entre populations.
Article à lire : La duplication des gènes, moteur de l’évolution, par Joseph Schacherer
La prédiction génétique à l'heure de la bio-informatique et du séquençage du génome => : https://view.genial.ly/6116761f9d2bd20dd5d32978 (Jeu sérieux sur la mucoviscidose)
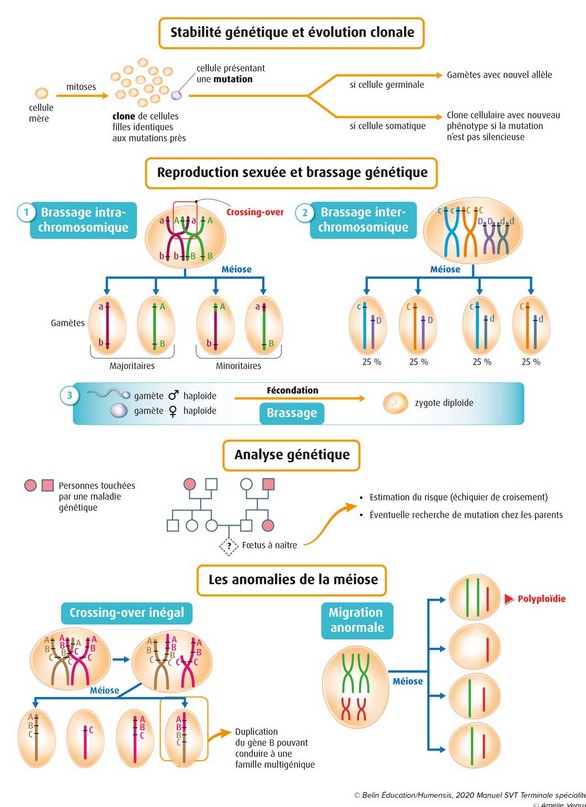
D'après
Belin (SVT Tle)